Journal Club: Sparrows show us a new way to have sexes
I’ve always loved white-throated sparrows, Zonotrichia albicollis. Not only are these handsome birds the sister species to my own dissertation bird, the white-crowned sparrow, Z. leucophrys, but I think they are among the most fascinating bird species in the world. In fact, I am so captivated by this species that the chance to pursue my “dream research project” on my beloved lories was all that stopped me from looking at the intricacies of white-throated sparrow evolution, ecology and behaviour.
Ever since white-throated sparrows were first discovered, it was known that they have two distinct colour polymorphisms (also known as “morphs”); some have black-and-white head stripes (“white-striped” or “WS”) and others have brown-and-tan head stripes (“tan-striped” or “TS”):
Comparison between white-throated sparrow colour morphs:
white-striped (WS) morph (left) and tan-striped (TS) morph (right).Image: Allison Reid. (NOTE: Permission for use with this article was granted by Dr Donna Maney.)
This colour variation corresponds perfectly to one of two alternative life history strategies (doi:10.1093/beheco/14.3.425). The more contrast-y WS males sing often and are more aggressive than their plainer TS brothers. WS males frequently intrude into neighboring males’ territories, they invest less time into guarding their mates, they provide almost no parental care to their chicks and, as you probably suspect, they are avid philanderers. They also have higher testosterone concentrations circulating in their bloodstreams. After learning these things, if you predicted that the flamboyant WS males are neither devoted mates nor caring fathers, well, you’d be correct.

A singing male white-throated sparrow, Zonotrichia albicollis (WS morph).
Visit TheMusicOfNature‘s YouTube channel.
But what’s good for the “gander” also works well for the “goose” in this monomorphic species: like WS males, the more contrast-y WS females sing often, are more aggressive and provide less parental care than their less showy TS sisters, and the WS females are — for want of a better description — raging nymphomaniacs: soliciting copulations from their mates twice as often as TS females. WS females also have higher testosterone levels than their TS sisters. Female WS morphs are the birdie equivalent of “Amazon women warriors” compared to the plainer TS females.
Predictably, these life history strategies also affect white-throated sparrows’ mating choices, where the old addage — “opposites attract” — applies. WS and TS individuals show an exceptionally strong negative assortative mating pattern: almost 100 percent of all breeding pairs of white-throated sparrows comprise individuals of opposite morphs. Perhaps recognising that the devoted and caring TS males are most desirable, female WS sparrows specifically choose them for their mates. If this is the case, the less aggressive TS females lose out and are stuck with the flamboyant but philandering WS males.
These reproductive choices have led to several interesting consequences that were uncovered by molecular techniques. First, when researchers used DNA fingerprinting techniques to identify the father of the sparrow chicks, they found that extrapair fertilisations were common for WS male-TS female pairs (comprising nearly 32 percent of their nestlings and more than 56 percent of their broods), whilst extrapair offspring were rare for TS male-WS female pairs (only 4.4 percent of nestlings and 6.3 percent of broods).
Researchers also looked to see if some sparrows were leaving eggs in their neighbors’ nests for them to care for. Some conspecific brood parasitism was discovered among TS male-WS female pairs (4.4 percent of nestlings, and 12.5 percent of broods), but no cases of brood parasitism were identified for WS male-TS female pairs — perhaps because the “homebody” TS females stayed in such close proximity to their nests that unwanted foster parenting was prevented.
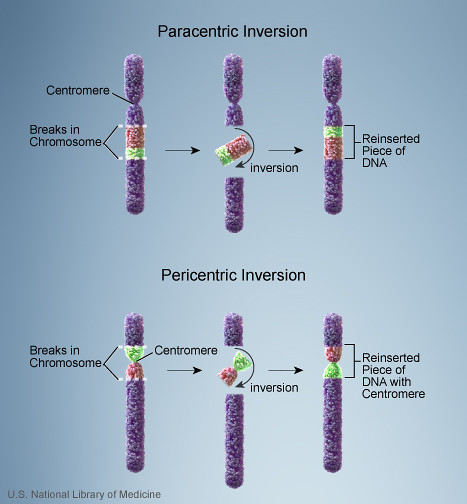
But what is the genetic foundation for these dramatic alternative life history strategies? Chromosomal staining methods revealed that white-throated sparrows have a very large mirror-image end-to-end chromosomal rearrangement: roughly 1000 genes on chromosome number 2 have flipped around (doi:10.1126/science.154.3756.1571). This type of rearrangement is known as a chromosomal inversion:
Image: National Library of Medicine/public domain.
This chromosomal inversion, known as the ZAL2 and ZAL2m genotype, is found with a 3:1 frequency within the population. Heterozygous (ZAL2/ZAL2m) birds are the showy WS morphs, whilst homozygous ZAL2/ZAL2 birds are the plainer TS morphs. These phenotypes occur at a 1:1 frequency among both males and females in the population, suggesting that it is not sex-linked and that each phenotype has particular selective advantages for both sexes. It is interesting to note in passing that ZAL2m homozygotes are vanishingly rare: after looking through the chromosomes of more than 600 white-throated sparrows, only one such individual has been identified so far.
But further research shows that the molecular behaviour of this chromosome is even more peculiar. A fascinating paper was just published by three people, including Donna Maney, Associate Professor of Psychology at Emory University and a former colleague of mine from graduate school, that reports on extreme linkage disequilibrium over the entire inverted region on chromosome 2 (doi:10.1038/hdy.2010.85). This means that the inverted region does not swap small genetic segments with its sister chromosome when egg and sperm cells are being created. This process, known as “crossing over”, is a normal part of meiosis:
Image: Thomas Hunt Morgan. (1916) A Critique of the Theory of Evolution, page 132 (public domain).
Basically, chromosome 2 acts like it is two different chromosomes, even though the inverted/non-inverted regions are stuck together on the same chromosome. This is really weird, but scientists have actually seen this before — this lack of crossing over, this linkage disequilibrium on the sparrows’ chromosome 2 is consistent with what we see for sex chromosomes. In fact, molecular evolutionary evidence indicates that the two sex chromosomes originally were one pair of autosomes — non-sex chromosomes — before evolving into separate X and Y chromosomes (or Z and W chromosomes in birds). Linkage disequilibrium was the critical event that had to occur before the sex chromosomes could evolve along separate pathways.
And evolve separately they did: in humans and other mammals, the X and Y chromosomes are dramatically different in size — X is one of the largest chromosomes and contains thousands of genes, whilst Y is the smallest and has only 70 or so genes — and they share almost none of their genes in common.
Yet despite their isolation, these two dramatically different chromosomes can still pair up during meiosis because they both retain a small region in common that allows them to recognise each other. That small region — the pseudoautosomal region (PAR) — behaves like an autosome because it still crosses over and thus, PAR has a shared evolutionary history between two otherwise separate and distinct sex chromosomes.
This separate evolutionary trajectory allows scientists to reconstruct the history of that chromosome. Since errors, or mutations, occur randomly and at a constant rate, the more generations of isolation that identical DNA regions experience, the greater their overall sequence divergence will be. Thus, evolutionary biologists can take advantage of this by analysing DNA sequence divergence and using it as a molecular clock to determine a chromosome’s age.
Based on these molecular analyses, we find that avian sex chromosomes, which arose around 310 million years ago, have a similar evolutionary history to mammalian sex chromosomes, which first appeared about 320 million years ago. Even though the avian system is somewhat different to mammals’ — for example, female birds are the heterogametic sex (ZW) whilst males are homogametic (ZZ), which is the opposite of mammals — the avian ZW sex-determining system shares many similarities with mammals: the Z chromosome is very large and gene-rich (similar to the mammalian X), whilst W is very small and contains few genes (like the mammalian Y). Also like mammalian XY, avian ZW have a PAR where crossing over occurs.
So even though white-throated sparrows already have sex chromosomes, they also have a second chromosomal system that behaves like sex chromosomes. Which leads me to ask: are white-throated sparrows evolving a second pair of sex chromosomes? If so, what might that system look like after its evolved further? The very idea boggles the mind and sounds like something you’d read about in science fiction. But this fascinating bird is alive and singing right in front of many North Americans, if we only took a moment to stop, look and learn some of the lessons that a common backyard bird has to teach us about evolution — and about life.

Sources:
Huynh, L., Maney, D., & Thomas, J. (2010). Chromosome-wide linkage disequilibrium caused by an inversion polymorphism in the white-throated sparrow (Zonotrichia albicollis). Heredity, 106 (4), 537-546 DOI: 10.1038/hdy.2010.85
Joron, M., & Whibley, A. (2010). Stripes, sex and sparrows: what processes underlie heteromorphic chromosome evolution? Heredity, 106 (4), 531-532 DOI: 10.1038/hdy.2010.106
Matsunaga, S. (2009). Junk DNA promotes sex chromosome evolution. Heredity, 102 (6), 525-526 DOI: 10.1038/hdy.2009.36
Moore, R. (2008). Sex chromosome evolution: A ‘missing link’ in the evolution of sex chromosomes. Heredity, 102 (3), 211-212 DOI: 10.1038/hdy.2008.115
Tuttle, E. (2003). Alternative reproductive strategies in the white-throated sparrow: behavioral and genetic evidence. Behavioral Ecology, 14 (3), 425-432 DOI: 10.1093/beheco/14.3.425
Thorneycroft, H. (1966). Chromosomal Polymorphism in the White-Throated Sparrow, Zonotrichia albicollis (Gmelin). Science, 154 (3756), 1571-1572 DOI: 10.1126/science.154.3756.1571
NOTE: this article is mirrored from Punctuated Equilibrium (hosted by The Guardian). Comments are not mirrored and are unique to the site where they were published.
That’s an amazing story. Thank you so much for summarizing what would have taken me weeks to tease apart. I took genetics as a college sophomore 21 years ago, and yet I still was able to follow your clear explanations. I wish I could come back in a million years to see what becomes of this species.