Journal Club: Jumping genes reveal deep relationships between parrots and songbirds
SUMMARY: A new study adds support to two earlier reports that songbirds and parrots are each other’s closest relatives (Psittacopasserae), indicating that vocal learning abilities appeared in this group of birds 30 million years earlier than originally assumed.
Passerines and parrots share a common ancestor as well as the ability to learn vocalization. Vocal learning may have evolved 30 million years earlier than previously thought.
Image: Corn bunting, Miliaria calandra. Kriegs/LWL (with permission) [velociraptorise] DOI: 10.1038/ncomms1448
{kind=link}
![]() Birds share many characteristics with humans. Unlike our fellow mammals, which learn about the world primarily by sniffing crotches, birds and humans have excellent colour sight, are highly auditory and some groups possess superior vocal learning abilities. Thus birds — especially songbirds and parrots — are the most important teachers that we have, particularly for scientists who study higher cognitive processes such as vocal learning and memory. Yet even though birds are the most studied group of animals in the world, the evolutionary origins of several major avian groups, particularly passerines and psittacines, remain shrouded in the mists of time.
Birds share many characteristics with humans. Unlike our fellow mammals, which learn about the world primarily by sniffing crotches, birds and humans have excellent colour sight, are highly auditory and some groups possess superior vocal learning abilities. Thus birds — especially songbirds and parrots — are the most important teachers that we have, particularly for scientists who study higher cognitive processes such as vocal learning and memory. Yet even though birds are the most studied group of animals in the world, the evolutionary origins of several major avian groups, particularly passerines and psittacines, remain shrouded in the mists of time.
Roughly 50 percent of all birds are passerines (songbirds), yet their relationships remain enigmatic. For example, morphological studies suggested that songbirds are a relatively young group within Neoaves (Neoaves includes all birds except the ratites — ostriches and similar — and the Galloanserae — “waterfowl”, “wildfowl” and “landfowl”), and that the sister group to the songbirds may be woodpeckers [doi:10.1111/j.1096-3642.2006.00293.x] and rollers [doi:10.1007/BF02101113] or cuckoos [doi:10.1046/j.1439-0469.2003.00230.x]. But mitochondrial studies indicate a number of conflicting relationships between passerines and the other Neoaves, and nuclear DNA analyses provide yet more conflicting data. Basically, none of our methodologies were providing clear information, so our understanding of songbird relationships were confused and confusing.
Then three years ago, a fascinating paper was published describing a DNA study that shook the foundations of our understanding of the avian tree of life. This study examined almost 32 kilobases of nuclear DNA sequences from 19 independent loci for 169 species representing all major extant avian groups — a tremendous number at the time. The most stunning revelation was that parrots and songbirds are sister taxa — each other’s closest living relatives. Was that proposed psittacine-passerine phylogenetic relationship real or was it artifact? Even though this finding agreed with an earlier study that proposed the same relationship [doi:10.1098/rsbl.2006.0523], this finding had the scientific and bird-watching communities abuzz. Everyone, it seemed, had an opinion about this.

The scarlet macaw, Ara macao, and other parrots learn vocalizations. Their closest living relatives, passerines, are also adept vocal learners, indicating that vocal learning may have evolved in their common ancestor.
Image: Kriegs/LWL (with permission) [velociraptorise] DOI: 10.1038/ncomms1448
Now there’s another study that adds yet more support to those original findings. Published by a team of German investigators led by Münster University graduate student Alexander Suh, this hot-off-the-presses research takes an even closer and more detailed look at the proposed relationship between parrots and songbirds by using yet another method: retroposons.
Retroposons are “jumping genes”. They are repetitive DNA fragments that insert randomly into the genome after being copied (“reverse transcribed”) from an RNA intermediary. Whilst the original repetitive retroposon sequence identities and locations are inherited like any other genetic loci, the new insertions (along with any changes in sequence) are unique and are reliably inherited from the time they are inserted. The resulting patterns of change are straightforward and can be traced through ensuing generations for more than 100 million years — major evolutionary timescales. In short, retroposons are molecular fossils.
Based on what we know of retroposons, it is reasonable to predict that they could be used as phylogenetic markers that may provide some intriguing insights into previously unresolved deep evolutionary relationships found early avian evolution, but no one was quite sure what Mr Suh and his team would actually discover.
There are more than 200,000 retroposed elements (REs) present in the chicken and zebra finch genomes, so Mr Suh and his team chose the two most numerous, comprising more than 97 percent of all avian REs: the chicken repeat 1 (CR1) family of long interspersed elements (LINEs) and the long terminal repeat elements (LTRs) of endogenous retroviruses.
Despite this huge number of loci, when the team analysed their chosen REs for bird-specific insertions in key avian groups, they identified several consistent patterns: (1) some retropsons (black balls) had a small deletion that was present in some avian CR1s, but was absent in all other avian and reptilian CR1s; (2) the presence or absence of specific RE markers (dark grey balls) are consistent with one another; and (3) some REs were inserted at the beginning of neoaves (light grey balls on grey gradient [label F]) and the RE dimorphisms were then subjected to incomplete lineage sorting. And the data were used to reconstruct an unambiguous family tree (figure 1):

Figure 1 | Retroposon evidence for the early branching events in the avian tree of life. [velociraptorise]
As you can see in the above phylogenetic tree, the retroposon evidence within Neoaves shows unmistakable presence/absence patterns. For example, the previously reported “landbird” assemblage (figure 1, branch G; two REs; also see doi:10.1098/rsbl.2006.0523) was recovered, as was a novel clade consisting of all “landbirds”, excluding mousebirds (figure 1, branch H; two REs; also see doi:10.1126/science.1157704) and these data also indicated a close affinity among seriemas, falcons, parrots and passerines (figure 1, branch I; two REs). Also note that these letters correspond to the labeled branches: A, Aves; B, Neognathae; D, Neoaves; F, incongruent markers; G, ‘landbirds’; H, ‘landbirds’ without mousebirds; I, Eufalconimorphae + seriemas; J, Eufalconimorphae; K, Psittacopasserae; L, Passeriformes.
In short, the most important take-away message from the above phylogram is that songbirds and parrots are sister taxa, and their closest relative is falcons (Falco) — and falcons are not closely related to the other raptors (hawks and eagles; Buteo). For the specialist, the parrot-passerine (Psittacopasserae) relationship is an important finding, because until very recently, parrots and songbirds were considered to be only distantly related.
The consistency between these markers surprised the team.
“I was very surprised that (after all these controversies between and within mitochrondrial, nuclear and DNA-hybridization datasets regarding the neoavian radiation) we actually recovered something that appears to be non-conflicting retroposon markers”, wrote Mr Suh in an email message.
But when did these retroposons do most of their “jumping”? The team looked for nested retroposons (sequence data not shown here); younger REs inserted inside older (inactive) REs that were already in place (figure 3):
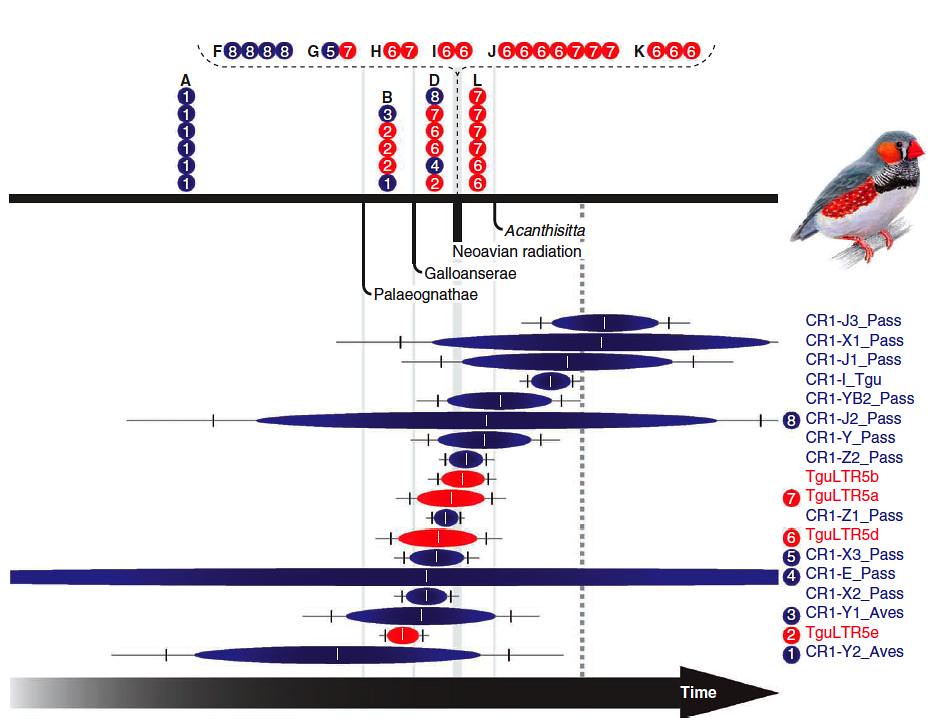
Figure 3 | Chronology of Mesozoic retroposon activity in the zebra finch genome. [velociraptorise]
The team examined 995 nested retroposons in the zebra finch genome (blue balls are CR1 or red balls are LTRs; numbers indicate specific RE subtypes and the horizonal blue or red lines indicate “activity periods”). They identified which retroposon markers were inserted during the neoavian radiation (dashed bracket) at specific times during avian evolution up until the estimated end of the Mesozoic Era at the Cretaceous/Tertiary boundary (grey dashed vertical line).
These divergences are denoted in the above figure, starting from the left side, as follows: the first pale grey vertical bar indicates when the Paleognaths (ratites; ostriches and similar birds) split away from the Neognaths (all the other birds); the second pale grey vertical bar indicates when the Galloanserae (“fowls”) split away from the Neoaves; the broader pale grey vertical bar corresponds to the rapid Neoavian radiation; and the pale grey vertical bar on the right side denotes when the Acanthisittidae split away from the oscines (Acanthisittidae are similar to suboscines because they do not learn their songs).
The most important thing to note in the chromogram above is that some RE subtypes were active during relatively short periods whilst others were not, yet these REs provide a surprisingly consistent estimate for retroposon nesting events during the Mesozoic evolution of birds.
Based on the topology of the phylogenetic tree (figure 1), we can see that vocal learning first appeared in the shared ancestor for parrots and songbirds — after the falcons diverged, as summarised below (figure 4):

Figure 4 | Evolution of vocal learning in birds. [velociraptorise]
This study underscores the striking neuroanatomical and gene expression similarities between parrots and songbirds, indicating that their vocal learning ability originally appeared in their shared ancestor 30 million years earlier than first predicted.
“With the strong retroposon support for Psittacopasserae (parrots + passerines), it seems plausible (and quite parsimonious) to assume that their similarities are not due to convergent evolution, but to shared ancestry”, wrote Mr Suh.
“Passerines and parrots are among those few avian groups that are capable of vocal learning and the new study suggests that this capability already originated in the last common ancestor of the group,” wrote Gerald Mayr, in email. Dr Mayr, a paleo-ornithologist and curator of ornithology at the Senckenberg Museum in Frankfurt, Germany, was not involved in the study.
“Many studies on vocal learning used either zebra finches [passerines] or parrots as model organisms, and recognition of close affinities between parrots and passerines may now make it much easier to directly compare and understand the data obtained from these different bird groups.”
Another interesting thing to note is that even though hummingbirds are also vocal learners, this study shows that they are only distantly related to Psittacopasserae, so their vocal learning capability evolved independently after they diverged from the swifts, their sister group, which lacks this ability.

The lesser kestrel, Falco naumanni, and all other falcons are not related to hawks, eagles and vultures. Instead, they are the closest living relatives of Psittacopasserae (passerines and parrots).
Image: Kriegs/LWL (with permission) [velociraptorise] DOI: 10.1038/ncomms1448
Apart from the fact that everyone is familiar with both parrots and songbirds, and is interested to learn more about them, this study also shows that analyses of “jumping genes” are a powerful new tool that can help to clarify evolutionary relationships among birds.
Even though the retroposon data is fully independent of nucleotide sequence analyses it still verified previous findings and provided robust support for some relationships, such as the Psittacopasserae, noted Mr Suh.
“If an understanding of the basic neural mechanisms of vocal learning is eased by recognition of close affinities between parrots and songbirds, this will perhaps also help to better understand vocal learning in humans,” Dr Mayr concluded.
Sources:
Suh, A., Paus, M., Kiefmann, M., Churakov, G., Franke, F., Brosius, J., Kriegs, J., & Schmitz, J. (2011). Mesozoic retroposons reveal parrots as the closest living relatives of passerine birds. Nature Communications, 2 DOI: 10.1038/ncomms1448
Alexander Suh [emails; 18 & 23 August 2011]
Gerald Mayr, curator of ornithology, Senckenberg Museum [email: 23 August 2011]
Background:
Here’s what I wrote about a Science paper that might be of interest regarding the deep phylogenetic relationships between birds.
Hackett, S., Kimball, R., Reddy, S., Bowie, R., Braun, E., Braun, M., Chojnowski, J., Cox, W., Han, K., Harshman, J., Huddleston, C., Marks, B., Miglia, K., Moore, W., Sheldon, F., Steadman, D., Witt, C., & Yuri, T. (2008). A Phylogenomic Study of Birds Reveals Their Evolutionary History. Science, 320 (5884), 1763-1768 DOI: 10.1126/science.1157704
Ericson, P., Anderson, C., Britton, T., Elzanowski, A., Johansson, U., Kallersjo, M., Ohlson, J., Parsons, T., Zuccon, D., & Mayr, G. (2006). Diversification of Neoaves: integration of molecular sequence data and fossils. Biology Letters, 2 (4), 543-547 DOI: 10.1098/rsbl.2006.0523
Universität Münster News aus dem Bereich Presse (Deutsch)
Funding for this research was provided by Deutsche Forschungsgemeinschaft (the German Research Foundation) and the Medizinische Fakultät der Westfälischen Wilhelms-Universität Münster (Medical Faculty of the Westphalian Wilhelms-University of Muenster).
.. .. .. .. .. .. .. .. .. .. .. ..
twitter: @GrrlScientist
facebook: grrlscientist
email: grrlscientist@gmail.com